Coelomycetes

A pycnidium of Phoma. Pycnidia are often spherical or inversely pearshaped (obpyriform) and their internal cavity is lined with conidiophores. When ripe, an opening generally appears at the top, through which the pycnidiospores escape. Pycnidia are not to be confused with perithecia which produce ascospores.
A class of conidial fungi where the conidia are formed within a cavity lined by fungal or host tissue.
The fruiting stuructres may be spherical with an apical openig (pynidia) or saucer shaped (acervuli). The production of conidia within a fruiting body distinguishes this group from the hyphomycetes which have "naked" conidia.
Pycnidia may be confused with ascocarps so its important to make the distinction between an acsospore and a conidium.
Species descriptions
-
Colletotrichum coccodes
Over 500 Colletotrichum species have been reported. C. coccodes is a common soil and plant pathogen widely distributed in Africa, Asia, Australasia, Europe, and the Americas. Colletotrichum species are a rare yet important cause of human mycotic keratitis (Hung et al., 2020).
RG-1 organism.

Sclerotia with setae of Colletotrichum coccodes.
Morphological description:
Colonies usually darkly pigmented with white aerial mycelium, consisting of numerous black sclerotia and light brown-coloured conidial masses, reverse is dark brown. Sclerotia are usually abundant, setose, spherical and are often confluent. Conidia are straight, fusiform, attenuated at the ends, 16-22 x 3-4 µm. Appressoria are common, clavate, brown, 11-16.5 x 6-9.5 µm, variable in shape.
Culture, conidia and appressoria of Colletotrichum coccodes.
Molecular identification: GAPDH gene sequencing is the recommended barcoding gene for species identification. ITS sequencing is not able to distinguish most species (Vieira et al., 2020).
Antifungal susceptibility: Colletotrichum species (Australian national data); MIC µg/mL. No ≤0.03 0.06 0.125 0.25 0.5 1 2 4 8 ≥16 AmB 15 1 3 5 3 2 1 ISAV 5 2 2 1 VORI 15 1 1 4 6 2 1 POSA 15 1 2 5 5 1 1 ITRA 15 1 2 2 3 1 6 References:
- Domsch, K.H., Gams, W. and Anderson, T.H. (2007) Compendium of soil fungi. Second Edition, IHW-Verlag, Germany.
- Hung, N., Hsiao, C.H., Yang, C.S., et al. (2020) Colletotrichum keratitis: A rare yet important fungal infection of human eyes. Mycoses, 63, 407-415.
- Vieira, W.A. dos S, Bezerra, P.A., Silva, A.C., et al. (2020) Optimal markers for the identification of Colletotrichum species, Molecular Phylogenetics and Evolution, 143, 106694-106694.
-
Lasiodiplodia theobromae
Synonymy:
Botryosphaeria rhodina; Botryodiplodia theobromaeLasiodiplodia theobromae is a well known plant pathogen reported from about 500 host plants, mainly confined to an area 40 degrees north to 40 degrees south of the equator. It has also been associated with ulcerated human cornea, lesions on nail and subcutaneous tissue.
RG-1 organism.

Lasiodiplodia theobromae pycnidia growing on carnation leaf agar.
Morphological description:
Colonies are greyish sepia to mouse grey to black, fluffy with abundant aerial mycelium; reverse fuscous to black. Pycnidia are simple or compound, often aggregated, stromatic, ostiolate, frequently setose, up to 5 mm wide. Conidiophores are hyaline, simple, sometimes septate, rarely branched cylindrical, arising from the inner layers of cells lining the pycnidial cavity. Conidiogenous cells are hyaline, simple, cylindrical to sub-obpyriform, holoblastic, annellidic. Conidia are initially unicellular, hyaline, granulose, sub-ovoid to ellipsoid-oblong, thick-walled, base truncate; mature conidia one-septate, cinnamon to fawn, often longitudinally striate, 20-30 x 10-15 µm. Paraphyses when present are hyaline, cylindrical, sometimes septate, up to 50 µm long.
Lasiodiplodia theobromae mature two-celled dark brown conidia with typical longitudinal striations.
Key features:
Coelomycete, with pycnidia producing characteristic two-celled, dark brown, striated conidia.Molecular identification:
Recommended genetic marker: EF-1α (de Hoog et al. 2015). ITS sequencing is useful for identifying clinically important species (Bagyalakshmi 2008).Antifungal susceptibility: Lasiodiplodia theobromae (Australian national data); MIC µg/mL No <0.016 0.03 0.06 0.125 0.25 0.5 1 2 4 8 16 32 ≥64 AmB 4 1 1 1 1 ISAV 1 1 VORI 4 1 2 1 POSA 3 1 2 ITRA 4 1 3 References:
- Bagyalakshmi, R., Therese, K.L., Prasanna, S., et al. (2008) Newer emerging pathogens of ocular non-sporulating moulds (NSM) identified by polymerase chain reaction (PCR)-based DNA sequencing technique targeting internal transcribed spacer (ITS) region. Current Eye Research, 33, 139-147.
- de Hoog, G.S., Guarro, J., Gene, J., et al. (2015) Atlas of Clinical Fungi (Version 4.1.2). Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands.
- Liu, J.-K., Phookamsak, R., Doilom, M., et al. (2012) Towards a natural classification of Botryosphaeriales. Fungal Diversity, 57, 149-210.
- Phillips, A.J., Alves, A., Abdollahzadeh, J., et al. (2013) The Botryosphaeriaceae: genera and species known from culture. Studies in Mycology, 76, 51-167.
-
Microsphaeropsis arundinis
Microsphaeropsis arundinis is an ascomycetous fungus that produces conidia within a pycnidium (an asexual fruiting body containing conidia). It is ubiquitous in soil and fresh water and typically inhabits terrestrial plant hosts. It typically inhabits terrestrial plant hosts and has a well-known association with Aruno donax, a garden escape weed known as ‘giant reed’ or ‘elephant grass’. M. arundinis is an emerging cause of phaeohyphomycosis in cats and immunosuppressed humans.
RG-1 organism.

Culture of Microsphaeropsis arundinis.
Morphological description:
Colonies growing slowly, with dense aerial mycelium, initially greenish-grey, later becoming dark brown to grey-brown. Hyphae are septate, pigmented, and irregularly shaped, with swollen segments up to 4 μm in diameter. Pycnidia are subspherical, 250-350 μm in diameter; with a pseudoparenchymatous wall composed of very densely packed cells that appear angular in cross section (textura angularis). Conidiogenous cells ampulliform, up to 5 μm long. Conidia brown, thick- and smooth-walled, cylindrical, 3.5-4.5 × 1.0-1.5 μm.
Microsphaeropsis arundinis showing pigmented septate hyphae, with swollen segments.
Key features:
Coelomycete, with ostiolate pycnidia, ampulliform conidiogenous cells, and small, smooth-walled, brown, cylindrical conidia.Molecular identification:
ITS and D1/D2 sequencing is recommended, especially as it may take many weeks before pycnidia are produced in culture (Reppas et al. 2015). Note: ITS and D1/D2 sequencing alone cannot discriminate between Microsphaeropsis arundinis and Paraconiothyrium cyclothyrioides, additional sequencing of ACT and b-tubulin is required (Gordon et al., 2012).References:
- de Hoog, G.S., Guarro, J., Gene, J., et al. (2015) Atlas of Clinical Fungi (Version 4.1.2). Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands.
- Gordon, R., Sutton, D., Thompson, E., et al. (2012) Cutaneous phaeohyphomycosis caused by Paraconiothyrium cyclothyrioides, Journal of Clinical Microbiology, 50, 3795-3798.
- Hall, M.R., Brumble, L.M. Mayes, M.A., et al. (2013) Cutaneous Microsphaeropsis arundinis infection initially interpreted as squamous cell carcinoma. International Journal of Dermatology, 52, 84-86.
- Kluger, E.K., Della Torre, P.K., Martin, P., et al. (2004) Concurrent Fusarium chlamydosporium and Microsphaeropsis arundinis infections in a cat. Journal of Feline Medicine and Surgery, 6, 271-277.
- Krockenberger, M.B., Martin, P., Halliday, C., et al. (2010) Localised Microsphaeropsis arundinis infection of the subcutis of a cat. Journal of Feline Medicine and Surgery, 12, 231-236.
- Pendle, S., Weeks, K., Priest, M., et al. (2004) Phaehyphomycotic soft tissue infections caused by the coelomycetous fungus Microsphaeropsis arundis. Journal of Clinical Microbiology, 42, 5315-5319.
- Reppas, G., Gottlieb, T., Krockenberger, M., et al. (2015) Microsphaeropsis arundinis an emerging cause of phaeohyphomycosis in cats and people. Microbiology Australia, 36, 74-78.
-
Neoscytalidium dimidiatum
Synonymy:
Hendersonula toruloidea; Scytalidium dimidiatum; Scytalidium hyalinum.Neoscytalidium dimidiatum is a coelomycete and recognised agent of onychomycosis and superficial skin infections, especially in tropical regions. The primary isolation of this fungus from clinical specimens may be difficult as isolates are sensitive to cycloheximide (actidione), which is commonly added to primary isolation media used for culturing skin scrapings.
The taxonomy of this species has been very confusing; the conidial state of S. dimidiatum was originally described under the name Hendersonula toruloidea. However it is phylogenetically remote from Scytalidium and has now been placed into Neoscytalidium (Crous et al. 2006, Machouart et al. 2012). Colourless mutants (previously known as Scytalidium hyalinum) often occur and have now been listed as variety N. dimidiatum var. hyalinum (Madrid et al. 2009).
Note:
Nattrassia mangiferae, previously considered the teleomorph form of S. dimidiatum, is now considered a distinct species, and has been placed in the genus Neofusicoccum, based on the lack of an arthoconidial anamorph (Crous et al. 2006, Machouart et al. 2012).RG-2 organism.

Microscopic morphology of the Neoscytalidium dimidiatum showing chains of one- to two-celled, darkly pigmented arthroconidia.
Morphological description:
Cultures are effuse, hairy, dark grey to blackish-brown, or white to greyish, with a cream-coloured to deep ochraceous-yellow reverse. Colourless mutants often occur. Arthroconidia are typically present in chains of one to two-cells, darkly pigmented, 3.5-5 x 6.5-12 µm, produced by the holothallic fragmentation of undifferentiated hyphae. Pycnidia, only occasionally formed in older cultures are black, ostiolate and contain numerous hyaline, flask-shaped phialides. Phialoconidia are at first one-celled and hyaline, later becoming three-celled, brown, with the centre cell darker than the end cells and are ovoid to ellipsoidal in shape.Molecular identification:
ITS and D1/D2 sequencing may be used for accurate species identification (Halliday et al. 2015).MALDI-TOF MS:
Alshawa et al. (2012) developed a spectral database for 12 different species of dermatophytes which also included Neoscytalidium dimidiatum and N. dimidiatum var. hyalinum. Correct identification of the species was obtained for 18/21 Neoscytalidium isolates (85.7%).Antifungal susceptibility: Neoscytalidium dimidiatum limited data (Australian national data); MIC µg/mL. Antifungal No ≤0.016 0.03 0.06 0.125 0.25 0.5 1 2 4 ≥8 AMB 12 1 1 3 5 2 VORI 12 1 2 1 1 3 1 3 POSA 12 1 1 2 1 1 1 5 ITRA 12 1 2 1 1 7 References:
- Alshawa, K., Beretti, J.L., Lacroix, C., et al. (2012) Successful identification of clinical dermatophyte and Neoscytalidium species by matrix-assisted laser desorption ionization-time of flight mass spectrometry. Journal of Clinical Microbiology, 50, 2277-2281.
- Crous, P.W., Slippers, B., Wingfield, M.J., et al. (2006) Phylogenetic lineages in the Botryosphaeriaceae. Studies in Mycology, 55, 235-253.
- de Hoog, G.S., Guarro, J., Gene, J., et al. (2015) Atlas of Clinical Fungi (Version 4.1.2). Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands.
- Frankel, D.H. and Rippon, J.W. (1989) Hendersonula toruloidea infection in man. Mycopathologia, 105,175-186.
- Halliday, C.L., Kidd, S.E., Sorrell, T.C., et al. (2015) Molecular diagnostic methods for invasive fungal disease: the horizon draws nearer? Pathology, 47, 257-269.
- Kidd, S., Halliday, C., Ellis, D. (2023) Descriptions of Medical Fungi (4th edition). CABI.
- Machouart, M., Menir, P., Helenon, R., et al. (2012) Scytalidium and scytalidiosis: what's new in 2012? Journal de Mycologie Medicale, 23, 40-46.
- Madrid, H., Ruíz-Cendoya, M., Cano, J., et al. (2009) Genotyping and in vitro antifungal susceptibility of Neoscytalidium dimidiatum isolates from different origins. International Journal of Antimicrobial Agents, 34, 351-354.
- McGinnis, M.R. (1980) Laboratory handbook of medical mycology. Academic Press, New York.
- Moore, M.K. (1986) Hendersonula toruloidea and Scytalidium hyalinum infections in London, England. Journal of Medical and Veterinary Mycology, 24, 219-230.
- Rippon, J.W. (1988) Medical mycology: the pathogenic fungi and the pathogenic actinomycetes, 3rd edition. W,B. Saunders Co, Philadelphia, USA.
- Sutton, B.C. and Dyko, B.J. (1989) Revision of Hendersonula. Mycological Research, 93, 466-488.
-
Phoma spp.
Members of the genus Phoma have a worldwide distribution and are ubiquitous in nature, with over 200 species having been described from soil, as saprophytes on various plants, and as pathogens to plants and humans.
RG-1 organism.
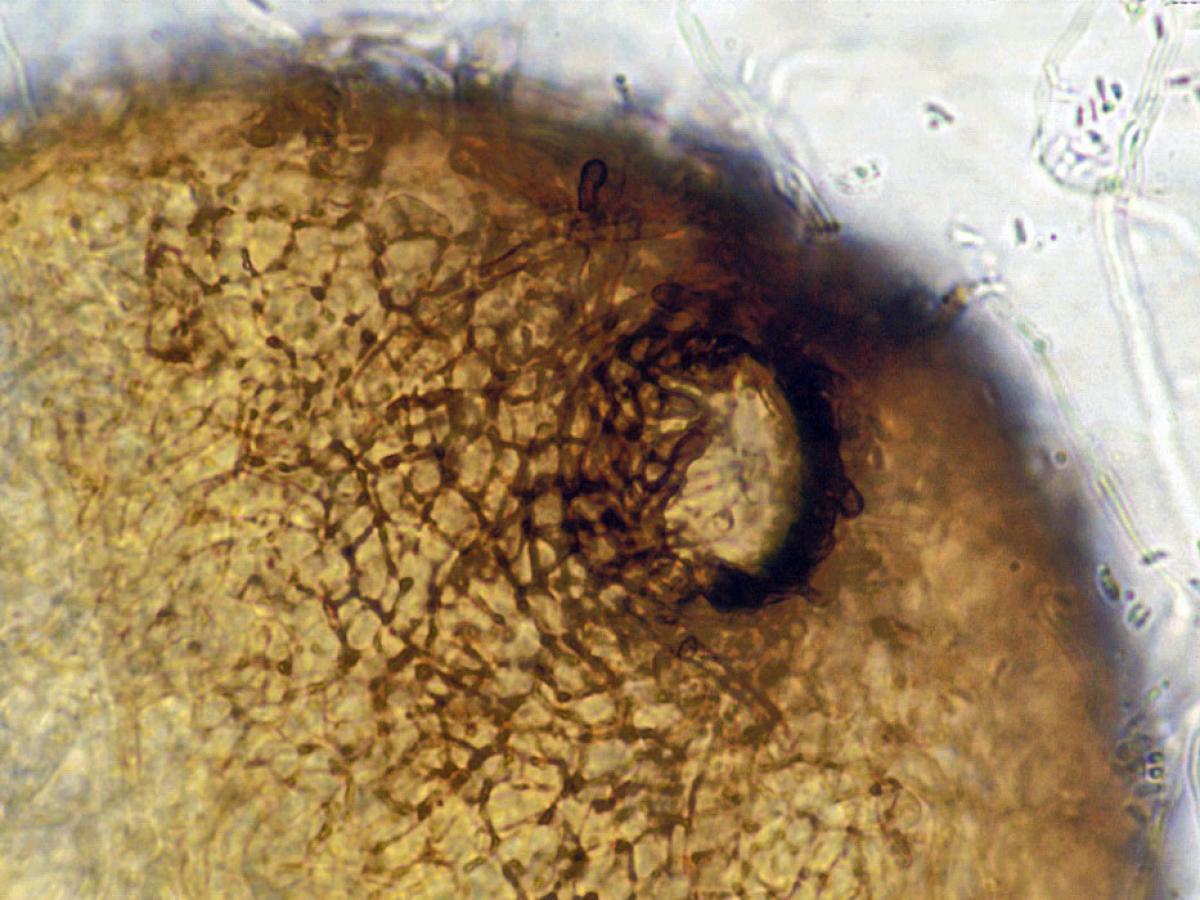
Pycnidium of Phoma.
Morphological description:
Colonies are spreading, greyish-brown, powdery or suede-like and produce large, globose, membranous to leathery, darkly pigmented, ostiolate pycnidia. Conidia are produced in abundance within the pycnidia on narrow thread-like phialides, which are hardly differentiated from the inner pycnidial wall cells. Conidia are globose to cylindrical, one-celled, hyaline, and are usually extruded in slimy masses from the apical ostiole.Molecular identification:
ITS, D1/D2, β-tubulin and 18S sequencing has been used to identify Phoma species (de Gruyter et al. 2009, Aveskamp et al. 2010). Note: Public sequence databases, particularly GenBank, contain many sequences from incorrectly identified species, making identifications of coelomycetous fungi very difficult, without confirmatory morphological studies.Key features:
Coelomycete, ostiolate pycnidia producing masses of slimy, hyaline, single-celled conidia.References:
- Aveskamp, M.M., de Gruyter, J., Woudenberg, J.H.C., et al. (2010) Highlights of the Didymellaceae: A polyphasic approach to characterise Phoma and related pleosporalean genera. Studies in Mycology, 65, 1-60.
- de Gruyter, J., Aveskamp, M.M., Woudenberg, J.H.C., et al. (2009) Molecular phylogeny of Phoma and allied anamorph genera: towards a reclassification of the Phoma complex. Mycological Research, 113, 508-519.
- de Hoog, G.S., Guarro, J., Gene, J., et al. (2015) Atlas of Clinical Fungi (Version 4.1.2). Centraalbureau voor Schimmelcultures, Utrecht, The Netherlands.
- Punithalingam, E. (1979) Sphaeropsidales in culture from humans. Nova Hedwigia, 31, 119-158.
- McGinnis, M.R. (1980) Laboratory handbook of medical mycology. Academic Press, New York.
- Montel, E., Bridge P.D., and Sutton, B.C. (1991) An integrated approach to Phoma systematics. Mycopathologia, 115, 89-103.
- Rippon, J.W. (1988) Medical mycology: the pathogenic fungi and the pathogenic actinomycetes, 3rd edition. W,B. Saunders Co, Philadelphia, USA.
- Samson, R.A., Hoekstra, E.S., Frisvad, J.C., et al. (1995) Introduction to food-borne fungi. Centraalbureau voor Schimmelcultures, P.O.Box 273, 3740 AG BAARN, The Netherlands.
- Sutton, B.C. (1980) The Coelomycetes, fungi imperfecti with pycnidia, acervuli and stromata. Commonwealth Mycology Institute, Kew, London.